Электромагнитная обработка модулирует газообмен во время прорастания семян гороха в условиях солевого стресса.
13. 04. 2026
1 Прогрессивные материалы и аддитивные технологии, Кабардино-Балкарский государственный университет им. Х.М. Бербекова, ул. Чернышевского, 123, Нальчик 360004, Россия
2 АО «Концерн «Гранит», Бульвар Гоголевский, 31, стр. 2, Москва 119019, Россия
3 Кафедра фундаментальных наук, Московский государственный технический университет им. Н.Э. Баумана, 2-я Бауманская ул., 5, Москва 105005, Россия
4 Федеральное государственное бюджетное научное учреждение «Федеральный научный центр овощеводства» (ФГБНУ ФНЦО), ул. Селекционная, 14, ВНИИССОК, Одинцовский район, Москва 143072, Россия
5 Институт агробиотехнологий им. А.В. Журавского, Коми научный центр, Уральское отделение, Российская академия наук, Ручейная ул., 27, Сыктывкар 167023, Россия
Аннотация
Электромагнитная обработка (ЭМП) может стимулировать прорастание семян и развитие растений, включая смягчение негативных последствий стрессовых факторов. Одним из неинвазивных подходов к обнаружению ранних эффектов воздействия ЭМП является изучение динамики газообмена на стадии набухания семян. Газовая хроматография использовалась для оценки влияния низкоинтенсивного нетермального ЭМП на концентрацию газов H2, O2, CO2 и NH3 в системе «почва – семена гороха» в оптимальных условиях и при солевом стрессе. Обработка ЭМП оказала зависящий от варианта эффект. В оптимальных условиях она стимулировала дыхание (концентрация O2 снизилась на 12%, CO2 увеличилась на 15%), тогда как при засолении концентрация обоих газов снизилась на 8−10% по сравнению с контролем. Выделение H2 оказалось чувствительным биохимическим маркером реакции на внешние факторы. В оптимальных условиях обработка ЭМП почти утроила выделение H2 и сместила его пик на один день раньше, что может указывать на ускоренную мобилизацию защитных систем семян при развивающейся гипоксии. Засоление снизило уровни H2 на порядок, в то время как обработка ЭМП стабилизировала скорость выделения H2, уменьшив ее почти вдвое. Таким образом, ЭМП следует рассматривать как модификатор метаболического ответа семян на условия набухания, а не только как стимулятор прорастания.
Ключевые слова: электромагнитный прайминг; солевой стресс; газообмен; ЭМП; водород; защита от стресса; биоактивация; гипоксия
Академический редактор: Франческо Маринелло
1. Введение
В последние годы предпосевной прайминг семян стал эффективной стратегией повышения толерантности растений к абиотическим стрессам [1]. Традиционно для этой цели использовались химические агенты (такие как дезинфектанты и составы для инкрустации). Однако химическая нагрузка на агроэкосистемы связана с экологическими рисками и рисками для здоровья и требует строгого контроля дозировок. В связи с этим все больше внимания уделяется разработке экологически безопасных («эко-дружественных») физических методов повышения стрессоустойчивости растений, которые могли бы стать альтернативой химическим регуляторам роста.
В контексте развития «зеленых» технологий особый интерес представляет метод предпосевной обработки семян с использованием низкоинтенсивных электромагнитных полей (ЭМП) как потенциально эффективный и экологически безопасный подход [2, 3].
Растения реагируют на геомагнитные поля (как сильные статические, так и переменные магнитные поля) многочисленными способами [4, 5]. Особое внимание уделяется биоэлектромагнитным взаимодействиям, лежащим в основе изменений метаболизма растений, стрессоустойчивости и продуктивности сельскохозяйственных культур, индуцированных импульсными магнитными полями [6]. Было установлено, что длительное воздействие слабого магнитного поля на растения может вызывать различные биологические эффекты на клеточном, тканевом и организменном уровнях [7]. В то же время подчеркивается ярко выраженная контекстная зависимость эффектов ЭМП, определяемая параметрами обработки, генотипом растений и условиями выращивания [8].
Большинство стратегически важных сельскохозяйственных культур, включая зернобобовые, подвержены воздействию засухи и засоления на ранних этапах онтогенеза как в природных экосистемах, так и в агроценозах [9, 10]. Засоление почвы выводит из традиционного сельскохозяйственного производства примерно три гектара пахотных земель каждую минуту [11]. На стадии прорастания солевой стресс индуцирует осмотический и ионный дисбаланс, повреждение мембран, ингибирование митохондриального дыхания, что проявляется в снижении поглощения O2 и выделения CO2, а также в нарушении окислительно-восстановительного гомеостаза [12-16]. В результате значительно снижаются темпы роста и уменьшается жизнеспособность проростков. Повышение толерантности сельскохозяйственных растений к абиотическим стрессам, таким как засоление, является одной из ключевых задач современной агроинженерии и растениеводства [17-19].
Согласно литературным данным, ЭМП может изменять проницаемость клеточных мембран, активность ферментов дыхательной цепи и антиоксидантной системы, а также повышать энергию прорастания и устойчивость растений к абиотическим стрессам, включая засоление и засуху [20-23]. Например, предпосевная обработка магнитным полем повысила засухоустойчивость сухих семян люцерны (Medicago scutellata L.) [22] и способствовала увеличению и стабилизации урожайности нута (Cicer arietinum L.) [24]. Обработка семян импульсным электрическим полем обеспечила значительное повышение скорости прорастания при воздействии 100–200mMNaCl (p≤0,05), при этом лучшие результаты прорастания (до 62,0±0,9%) наблюдались при энергии воздействия 17,28 Дж [21]. Электромагнитная стимуляция и солевой прайминг также доказали свою эффективность в повышении энергии прорастания и физиологической активности семян пшеницы в условиях богарного земледелия [25]. Предварительная обработка семян томатов переменным электромагнитным полем частотой 60Гц и магнитной индукцией 2–4мТл в течение 9мин привела к значительному увеличению процента и скорости прорастания, а также смягчила негативное влияние солевого стресса на фотосинтез и рост проростков [26]. В совокупности эти данные указывают на высокий потенциал ЭМП как инструмента для повышения толерантности растений к засолению.
Однако предыдущие исследования были в основном сосредоточены только на регистрации морфофизиологических и биохимических параметров проросших семян и развитых проростков, тем самым документируя лишь долгосрочные эффекты воздействия ЭМП. Напротив, критическая стадия, на которой семена переходят из жизнеспособного в активное метаболическое состояние, а именно динамика метаболической активации во время набухания семян сразу после обработки ЭМП, часто упускалась из виду. По большей части эта стадия развития характеризовалась исключительно сравнением скорости поглощения воды набухающими семенами между контрольной и обработанной группами [27]. Это связано с тем, что проведение детальных биохимических анализов активности требует нарушения целостности семян. Неинвазивным подходом, позволяющим косвенно проникнуть в суть процессов, происходящих в семенах во время набухания, является изучение динамики газообмена (поглощение O2, выделение CO2, H2 и NH3) [7, 28, 29].
Газообмен служит интегральным показателем метаболического состояния биологической системы и отражает активность митохондриального дыхания, окислительно-восстановительные процессы и адаптивные реакции растительного организма на стрессовые факторы [25, 26, 30, 31]. Изменения уровней CO2 и O2 коррелируют с интенсивностью аэробного дыхания через растительный цикл Кребса, скорость которого зависит от условий окружающей среды [7, 32]. Выделение H2 не связано напрямую с циклом Кребса и указывает на активацию альтернативных путей энергетического метаболизма, характерных для окислительно-восстановительного дисбаланса и гипоксических условий. В частности, его выделение может быть связано с активностью гидрогеназ и азотфиксирующих ферментов [33-35]. Молекулярный водород H2 также все чаще рассматривается как сигнальная молекула, участвующая в стрессовых реакциях растений. Сообщалось о положительном влиянии эндогенного и экзогенного H2 на стрессоустойчивость некоторых культур (таких как рис и рапс) к экологическим стрессорам [34, 36-38]. Основным источником NH3 в биологических системах считается дезаминирование аминокислот. В стрессовых условиях может активироваться как деградация белков, так и активность глутаматдегидрогеназы; этот фермент катализирует обратимую реакцию с участием α-кетоглутарата (промежуточного продукта цикла Кребса) [26, 28]. То есть у растений в условиях стресса субстраты цикла Кребса могут переключаться на удовлетворение других потребностей, например, на продукцию NH3. Однако мы не обнаружили в литературе исследований, в которых всесторонне анализировалась бы динамика выбросов ключевых газов в системе «почва – семена гороха», особенно в период набухания под влиянием обработки ЭМП, солевого стресса и их комбинированного действия.
Целью данного исследования была комплексная оценка влияния предпосевной обработки семян гороха ЭМП на динамику газообмена в условиях индуцированного засоления субстрата. Таким образом, задачи заключаются в следующем:
Количественно оценить вклад основных факторов («Семена», «Соль», «ЭМП») и их взаимодействий в изменения концентраций H2, O2, CO2 и NH3; Обсудить возможные физиолого-биохимические механизмы, лежащие в основе наблюдаемых изменений; Оценить потенциал использования ЭМП в качестве метода биостимуляции стрессоустойчивости.
2. Материалы и методы
Материалы: Почва, взятая в Чегемском районе (Кабардино-Балкарская Республика, Россия); поваренная соль (NaCl); семена гороха (Pisum sativum L.) сорта «Немчиновский 50», выведенного Федеральным государственным бюджетным научным учреждением «Федеральный исследовательский центр «НЕМЧИНОВКА», Одинцово, Московская область, Российская Федерация; (год репродукции 2022, партия N∘705), с влажностью 9−11%.
Оборудование: Генератор ЭМП, устройство «ТОР», произведенное АО «Концерн «Гранит» (Москва, Российская Федерация) (Рисунок 1). Режим обработки был настроен в соответствии с параметрами, рекомендованными авторами (Патент РФ N∘2765973 от 7 февраля 2022 г.). Габаритные размеры устройства: 205,4×185,1×432 мм; масса без упаковки: 8,5 кг; рабочая мощность: 12 Вт в рабочем диапазоне температур от −20∘C до +40∘C (потребляемая мощность в режиме охлаждения при температуре окружающей среды выше 40∘C: 150 Вт; напряжение: 220 В. Принцип работы устройства основан на применении специфического широкополосного, неионизирующего, нетермального импульсного электромагнитного излучения.
Рисунок 1.** Устройство «ТОР»

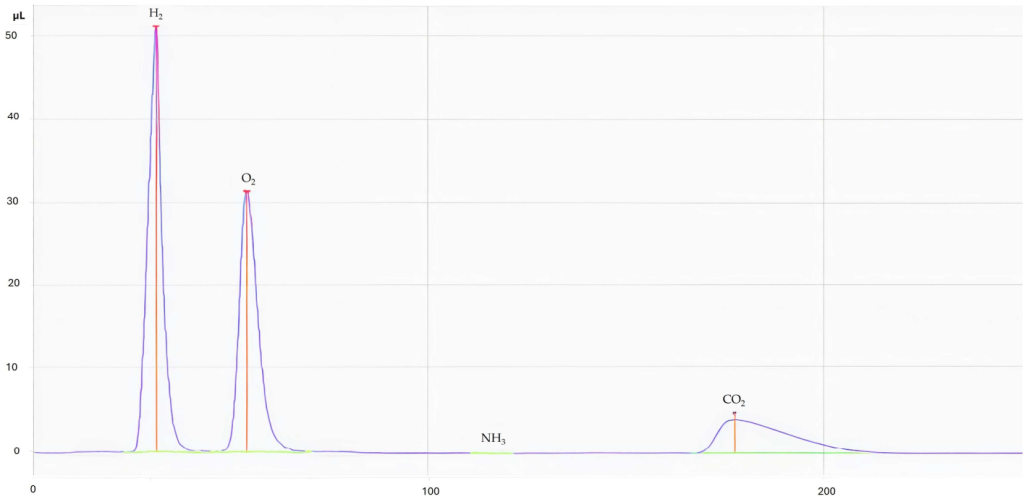
Использовался газовый хроматограф «ЦВЕТ-800» (ООО «ЦВЕТ», Дзержинск, Нижегородская область), оснащенный насадочной колонкой для разделения газов H2, O2, CO2 и NH3. Абсолютная калибровка для количественных расчетов была выполнена для всех газов с использованием чистых компонентов. Обработка данных проводилась с помощью программного обеспечения «Цвет-Аналитик v 1.03 E». Типичная хроматограмма для четырех газов представлена на Рисунке 2.
**Рисунок 2.** Типичная хроматограмма для четырех газов:
Характеристика почвы. Почва, использованная в исследовании, представляет собой чернозем выщелоченный оподзоленный, тип почвы, широко распространенный в речных долинах и предгорьях Чегемского района Кабардино-Балкарской Республики, Российская Федерация. Агрохимическая оценка образца почвы показала, что содержание основных элементов питания было типичным для данной географической зоны [39]. pH водной вытяжки составил 7,4 (ГОСТ 26483-85); содержание гумуса – 5,45% (метод Тюрина, ГОСТ 26213-91); щелочногидролизуемый азот – 100 мг/кг (метод Корнфилда) [40]; доступный фосфор (подвижные формы) – 35,0 мг/кг (метод Чирикова, ГОСТ 6204-91); обменный калий – 436,7 мг/кг (метод Чирикова, ГОСТ 6204-01).
Методика. Был рассчитан объем воды, который необходимо добавить к почвенному субстрату для достижения конечной влажности 18% (определено согласно ГОСТ 28268-89). Было установлено, что на 100 г почвенного субстрата необходимо добавить 40 мл воды. Партия семян гороха (всхожесть 99%) была откалибрована по размеру, и была отобрана фракция семян с однородной массой (0,200±0,002 г).
Дизайн эксперимента: Комбинации компонентов для газового анализа
Следующие комбинации изучаемых компонентов (почва, солевой раствор и семена гороха) были помещены в герметичные флаконы с септами для анализа газового состава (Таблица 1, Рисунок 3):
«Почва» – 10 г почвенного субстрата +4 мл дистиллированной воды;
«Почва + NaCl» – 10 г почвенного субстрата +4 мл 2,5% водного раствора NaCl;
«Почва + Семена» – 10 г почвенного субстрата +0,6 г семян гороха (три семени);
«Почва + NaCl + Семена» – 10 г почвенного субстрата +4 мл 2,5% водного раствора NaCl +0,6 г семян гороха (три семени).
1. Общая схема экспериментальных вариантов (проведено пять независимых экспериментов).
| Вариант | Компоненты системы | |||||
| Контроль (Необработанные) | Эксперимент (Обработанные ЭМП) | |||||
| Почва | NaCl | Семена | Почва | NaCl | Семена | |
| «Почва» | + | – | – | + | – | – |
| «Почва + NaCl» | + | + | – | + | + | – |
| «Почва + Семена» | + | – | + | + | – | + |
| «Почва + NaCl + Семена» | + | + | + | + | + | + |
**Методика: Оценка влияния ЭМП-прайминга на газообмен семян гороха в условиях солевого стресса**
Для каждой комбинации было подготовлено шесть герметичных флаконов с септами (4 комбинации ×6=24 флакона), которые были разделены на две группы (4 комбинации ×3=12 флаконов в группе). Одна группа была обработана ЭМП с использованием устройства «ТОР» (экспериментальная группа) сразу после герметизации в герметичных флаконах с септами, с временем экспозиции 5 минут и расстоянием два метра от излучателя до объекта. Вторая группа состояла из необработанных контрольных флаконов (контрольная группа). Всего эксперимент включал 8 вариантов, каждый с трехкратной повторностью. Полная схема эксперимента представлена в Таблице 1.
Лабораторный эксперимент проводился при рассеянном естественном дневном свете при комнатной температуре (+22−24∘C). Отбор проб газа проводился ежедневно из герметичных флаконов с септами путем введения шприца через септу и отбора 1 мл газовой смеси; пробы отбирались в утренние часы и вводились в газовый хроматограф для количественного определения целевых газов в течение пяти дней. С помощью программного обеспечения «Цвет-Аналитик v 1.03 E» регистрировалось количество каждого газа (в μЛ) на типичной хроматограмме для четырех газов (Рисунок 3).
Эксперимент был повторен в пяти независимых сериях, которые дали сопоставимые результаты. Анализ всего набора данных и усредненных параметров из пяти независимых экспериментов был выполнен с использованием дисперсионного анализа (ANOVA) для оценки значимости факторов и их взаимодействий, а также регрессионного анализа. Обработка данных проводилась с использованием программного обеспечения Statistica 10.0 и R 4.2.1. Для более детального анализа динамики процессов рассчитывалась скорость изменения концентрации каждого газа (ΔC/Δt) за последовательные дневные интервалы (1→2,2→3,3→4,4→5). Скорость рассчитывалась как разница концентраций между последующим и предыдущим днем для каждой отдельной повторности; после этого вычислялось среднее значение и стандартная ошибка для каждой комбинации факторов. Статистический анализ скоростей проводился с использованием трехфакторного дисперсионного анализа (факторы: вариант системы, временной интервал, условие, ЭМП/контроль) с последующим апостериорным критерием Дункана для выявления значимых различий между контрольной и экспериментальной группами в конкретных условиях [41].
3. Результаты
Изучение взаимодействий между абиотическими стрессорами и физическими методами воздействия на растения представляет значительный интерес в современной агробиологии. В данном исследовании анализируется комбинированное влияние NaCl и обработки ЭМП на динамику газообмена (O2, CO2, H2, NH3) в системе «почва/засоление/прорастающие семена гороха». Для количественной оценки интенсивности процессов газообмена также были рассчитаны скорости изменения концентраций газов (Δ[Газ]/Δt,μЛ/день) с использованием метода конечных разностей на интервалах между измерениями. Этот подход позволил выявить критические периоды максимальной метаболической активности прорастающих семян гороха и оценить динамическое влияние факторов (NaCl, ЭМП).
Динамика O2/CO2O2/CO2 (Рисунки 4 и 5). В отсутствие семян концентрация O2 оставалась постоянно высокой, в то время как выделение постепенно увеличивалось за счет активности почвенной микробиоты. ЭМП значительно стимулировало микробное дыхание, что проявлялось в постоянно более высоких концентрациях CO2 во всех обработанных вариантах. Засоление («Почва + NaCl») ингибировало этот процесс, снижая потребление на и выделение на к 5-му дню по сравнению с незасоленными вариантами.
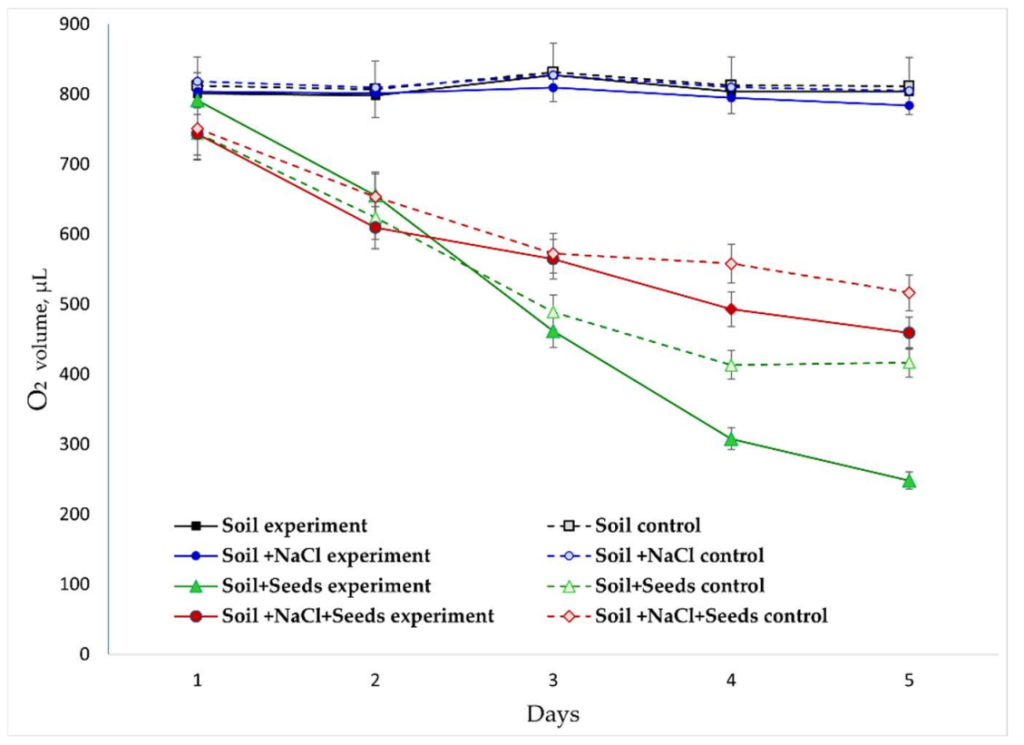
**Рисунок 4.** Динамика изменения концентрации O2 в газовой среде в разных экспериментальных вариантах.
Присутствие прорастающих семян в системе принципиально изменяет газообмен. Динамика потребления кислорода и выделения углекислого газа отражает интенсивность их дыхательного метаболизма, где наблюдается ожидаемое резкое снижение концентрации и увеличение , коррелирующее с активацией аэробного дыхания в семенах. Были идентифицированы две ключевые фазы дыхательной активности проростков: (1) фаза интенсивного потребления (дни 1-3) с пиковым потреблением (до −150 μЛ/день) и выделением (до +80 μЛ/день), особенно в варианте с обработкой ЭМП; (2) фаза стабилизации (дни 4-5), когда скорости газообмена и снижались, совпадая с фазой метаболической активации и началом роста корешка.
**Рисунок 5.** Динамика изменения концентрации CO2 в газовой среде в разных экспериментальных вариантах.
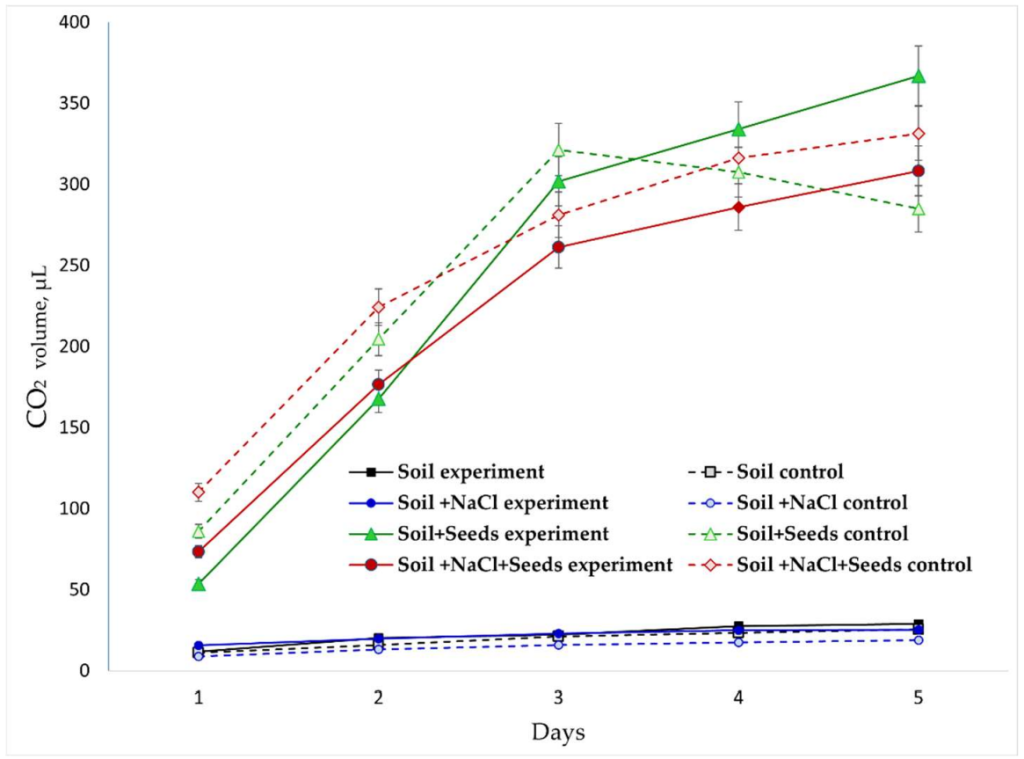
Обработка ЭМП оказала зависящий от варианта эффект: в варианте «Почва + Семена» она стимулировала дыхание (дополнительное снижение на 12%, увеличение на 15%). И наоборот, в стрессовом варианте «Почва + NaCl + Семена» эффект был противоположным: потребление было менее интенсивным, а выделение было на 8−10% ниже по сравнению с контролем в варианте «Необработанная почва».
Расчеты скорости изменения в варианте «Почва + NaCl + Семена» также указывают на защитное действие ЭМП. Ежедневное снижение потребления в обработанной группе было на 20−30% ниже, чем в контроле. Аналогично, увеличение концентрации под воздействием ЭМП было более постепенным (на ниже пиковых значений), что указывает на снижение окислительного «стрессового» всплеска (Таблица 2).
Расчет отношения скоростей выделения к скоростям потребления – аналога дыхательного коэффициента, – выявил следующие закономерности: В незасоленных условиях в контрольном варианте «Почва + Семена» что характерно для окисления углеводов. В ответ на ионный стресс в контроле увеличился до 1,3-1,5, что может указывать на вовлечение органических кислот в дыхание или частичную ферментацию дыхательного субстрата. В условиях засоления обработка ЭМП поддерживала значение близким к 1,0-1,2, что предполагает частичную компенсацию ингибирования дыхания набухающих семян и, вероятно, улучшенную энергетическую эффективность.
Таблица 2. Средняя скорость изменения концентрации газа (μЛ/день), отрицательные значения указывают на потребление) и результаты критерия Дункана для значимых интервалов.
| Система Вариант | Интервал (День № → День №) | Контроль (Среднее ± SE) | Эксперимент (ЭМП) (Среднее ± SE) | p-Уровень |
|---|---|---|---|---|
| ΔO₂/Δt | ||||
| Почва + Семена | 3 → 4 | -75,6 ± 3,2 a | -153,6 ± 5,1 b | <0,001 |
| 4 → 5 | -5,2 ± 2,1 a | -59,8 ± 4,3 b | <0,001 | |
| Почва + NaCl + Семена | 2 → 3 | -45,0 ± 2,8 a | -60,8 ± 4,5 b | 0,008 |
| 4 → 5 | -34,2 ± 1,9 a | -36,0 ± 3,2 b | 0,654 | |
| Почва + NaCl | 4 → 5 | -3,6 ± 0,9 a | -10,6 ± 1,2 b | <0,001 |
| ΔCO₂/Δt | ||||
| Почва | 4 → 5 | 2,08 ± 0,45 a | 1,40 ± 0,21 b | 0,022 |
| Почва + NaCl | 1 → 2 | 3,90 ± 0,32 a | 4,16 ± 0,28 b | 0,049 |
| 4 → 5 | 1,54 ± 0,15 a | 0,38 ± 0,09 b | <0,001 | |
| Почва + Семена | 1 → 2 | 118,6 ± 4,2 a | 114,1 ± 3,8 b | 0,047 |
| 4 → 5 | -22,6 ± 1,8 b | 32,8 ± 2,1 a | <0,001 | |
| Почва + NaCl + Семена | 2 → 3 | 60,8 ± 2,1 a | 76,8 ± 3,4 b | <0,001 |
| 4 → 5 | 15,4 ± 1,2 a | 22,4 ± 1,5 b | 0,001 | |
| ΔH₂/Δt | ||||
| Почва + Семена | 3 → 4 | 15,82 ± 2,25 a | 52,66 ± 3,25 b | <0,001 |
| 4 → 5 | 9,40 ± 1,85 a | 37,40 ± 2,15 b | <0,001 | |
| Почва + NaCl + Семена | 4 → 5 | 5,40 ± 0,35 a | 2,98 ± 0,25 b | <0,001 |
| Почва + NaCl | 4 → 5 | 0,0032 ± 0,0005 a | 0,0004 ± 0,0001 b | <0,001 |
a,bАпостериорный критерий Дункана использовался для сравнения средних значений между контролем и ЭМП в каждой комбинации обработка день. В таблице показаны средние значения в варианте; a и b указывают на достоверность различий, где значения с одинаковой буквой указывают на отсутствие значимого различия с вероятностью 95% согласно критерию Дункана [41].
Динамика H₂ (Рисунок 6). Выделение H₂ было обнаружено в основном в вариантах с прорастающими семенами, что подтверждает его биогенное происхождение. Максимальные значения (до 120 мкл) были зарегистрированы в варианте «Почва + Семена» при обработке ЭМП, где самая высокая скорость выделения H₂ наблюдалась на 3–4-й дни, достигая в среднем +52,7 мкл/день, что более чем в три раза превышало контроль (Таблица 2). Это соответствует фазе начальной активации роста зародышевого корешка и максимальной индукции активности гидрогеназы. Засоление («Почва + NaCl + Семена») послужило мощным фактором дистресса, снизив уровни H₂ на порядок. Скорости выделения H₂ были соответственно существенно ниже (<5,5 мкл/день), и их пик сместился на 4–5-й дни (Таблица 2). В отличие от абсолютных концентраций, анализ скоростей ясно показал, что обработка ЭМП в отсутствие солевого стресса не только увеличила общее выделение H₂, но и сместила его пик на более раннее время (с 4–5-х дней на 3–4-й), что может указывать на ускоренную мобилизацию защитных систем. В стрессовых условиях ЭМП стабилизировало выделение H₂ и уменьшило его почти вдвое, предотвращая резкие колебания (Таблица 2). Следует отметить, что вклад микроорганизмов в выделение водорода в нашем эксперименте был незначительным, и в варианте «Необработанная почва» выделений H₂ не было обнаружено вовсе.
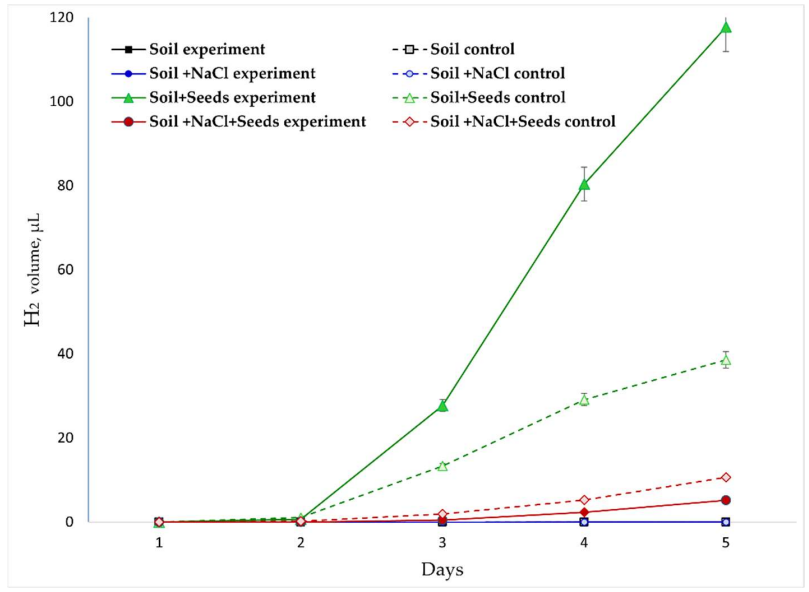
Рисунок 6. Динамика изменения концентрации H₂ в газовой среде в разных экспериментальных вариантах.
Динамика NH₃ (Рисунок 7). Концентрация NH₃ демонстрировала волнообразную динамику (5–9 мкл) без систематического влияния основных факторов в ходе краткосрочного эксперимента. Анализ скоростей подтвердил двухфазную динамику с положительными пиками на 2-й и 4-й дни (до +1,5 мкл/день) и отрицательной фазой на 3-й день (потребление до –1,0 мкл/день). Ни один из внешних факторов («Соль», «ЭМП») не оказал статистически значимого влияния на амплитуду этих пиков, лишь изменив их временную синхронизацию в пределах 0,5 дня. Присутствие прорастающих семян в анализируемых системах не изменило значимо уровень NH₃, что позволяет предположить, что его источником является преимущественно почвенная микробиота.
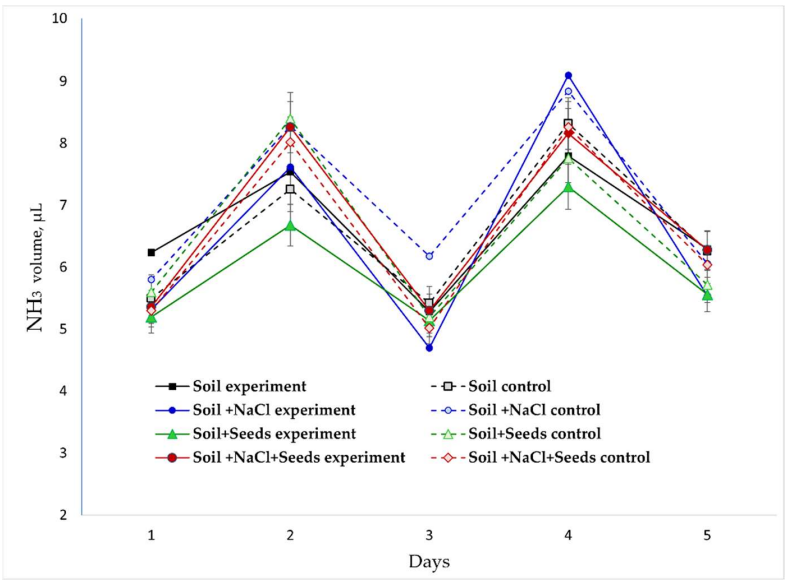
Рисунок 7. Динамика изменения концентрации NH₃ в газовой среде в разных экспериментальных вариантах.
Многофакторный дисперсионный анализ подтвердил статистически значимое влияние всех трех основных факторов и их взаимодействий на концентрации ключевых газов (p < 0,05) (Таблица 3). Доминирующим фактором было присутствие семян гороха, на которое приходилось примерно 60–70% общей изменчивости показателей O₂ и CO₂ и около 55% изменчивости H₂. Солевой стресс вносил вклад на уровне 8–12%, в первую очередь через взаимодействие с биологическим фактором, подавляя метаболическую активность. Обработка ЭМП объясняла 6–15% общей вариации, причем ее эффект был разнонаправленным, что выявило антагонистический характер влияния NaCl и ЭМП на метаболизм семян. В оптимальных условиях ЭМП действовало как стимулятор, проявляя синергизм с фактором семян; при засолении его роль смещалась в сторону защитной. Здесь обработка ЭМП продемонстрировала антагонизм по отношению к индуцированным стрессом процессам, уравновешивая солевой дистресс «ЭМП-эустрессом». Взаимодействия между факторами были статистически значимы и объясняли до 25% отклонений от аддитивной модели. Никакого значимого влияния факторов на эмиссию NH₃ обнаружено не было, что означает, что аммиак образуется более стабильно и, вероятно, контролируется фоновыми микробными процессами и равновесием NH₄⁺/NH₃.
Таблица 3. Доли влияния факторов и их взаимодействия на вариацию концентрации газа в системе (в среднем за всю совокупность данных).
| Фактор | O₂ | CO₂ | NH₃ | H₂ |
|---|---|---|---|---|
| Семена | 62 | 68 | 28 | 55 |
| NaCl | 7 | 12 | 9 | 8 |
| ЭМП | 11 | 15 | 6 | 12 |
| Семена × NaCl | 4 | 5 | 8 | 5 |
| Семена × ЭМП | 9 | 9 | 12 | 13 |
| NaCl × ЭМП | 3 | 4 | 5 | 4 |
| Семена × NaCl × ЭМП | 4 | 3 | 3 | 3 |
Был проведен комплексный корреляционный анализ для выявления взаимосвязей между всеми изучаемыми параметрами, включая бинарные факторы (Семена, NaCl и ЭМП). Этот анализ подтвердил тесную связь между дыхательными процессами; сильные отрицательные корреляции были обнаружены между концентрациями O₂ и CO₂ во всех вариантах, содержащих семена (коэффициент корреляции r от –0,85 до –0,95, p < 0,001). В присутствии ЭМП эти корреляции сохранялись, но с измененными наклонами регрессии, что указывает на модификацию эффективности дыхания. Влияние ЭМП было менее выраженным, но все еще значимым в вариантах без семян, особенно в отношении CO₂. Положительная корреляция была выявлена между выделением H₂ и уровнями CO₂ в контрольной группе (r ≈ 0,75–0,85), которая ослабевала под влиянием ЭМП (r ≈ 0,4–0,6). Это позволяет предположить, что ЭМП может разобщать связанные процессы анаэробного дыхания и ферментации, ведущие к образованию H₂. Никаких значимых корреляций не было обнаружено между NH₃ и другими газами, а также между параметрами в вариантах без семян.
Для выявления взаимосвязей между всеми изучаемыми параметрами на основе объединенного набора данных была рассчитана общая корреляционная матрица. Факторы «Семена», «NaCl» и «ЭМП» были закодированы как бинарные переменные (1 — присутствует, 0 — отсутствует). Полученные коэффициенты корреляции Пирсона представлены в Таблице 4, которая также подтверждает, что ключевым фактором, в первую очередь определяющим интенсивность газообмена, является биологическая активность прорастающих семян (сильные корреляции с концентрациями O₂, CO₂ и H₂). Более того, положительная корреляция между CO₂ и H₂ (r = 0,78) отражает активацию анаэробного метаболизма в условиях гипоксии, связанной с интенсивным дыханием (Таблица 4).
Таблица 4. Матрица коэффициентов корреляции Пирсона между экспериментальными факторами (наличие Семян, NaCl, ЭМП) и концентрациями газов (между всеми изучаемыми параметрами, включая бинарные факторы).
| Параметр | Семена | NaCl | ЭМП | O₂ | CO₂ | NH₃ |
|---|---|---|---|---|---|---|
| O₂ | -0,75 | 0,11 | -0,15 | — | — | — |
| CO₂ | 0,72 | 0,09 | 0,18 | -0,87 | — | — |
| H₂ | 0,69 | 0,21 | -0,25 | -0,65 | 0,78 | -0,08 |
| NH₃ | — | — | — | — | — | — |
Примечание: Коэффициенты, выделенные жирным шрифтом, значимы при p < 0,01. Для H₂ корреляции рассчитаны на основе данных за 2, 4 и 5-й дни.*
Влияние электромагнитного излучения на газообмен, хотя и будучи статистически значимым в целом (p < 0,001), в значительной степени зависело от состава системы и времени воздействия, что отражалось в величине коэффициентов корреляции (Таблица 4). ЭМП не оказывало независимого значимого влияния на газовый состав в абиотической почве. Однако в присутствии семян и в условиях солевого стресса оно действовало как метаболический модулятор, проявляя двойной физиологический эффект: стимулирующий в оптимальных условиях (усиление дыхания и выделения H₂) и защитный в условиях солевого стресса (метаболическая стабилизация и ингибирование выделения H₂ в сочетании со сниженным стрессовым дыханием).
4. Обсуждение
Неионизирующее электромагнитное воздействие служит значимым внешним регулятором скорости и направления ключевых метаболических процессов в системе «почва + прорастающее семя». Его эффекты зависят от системы и времени, наиболее ярко проявляясь при развивающейся кислородной недостаточности, прежде всего влияя на пути дыхательного метаболизма и анаэробную ферментацию набухающих семян. Высокая отрицательная корреляция между O₂ и CO₂ (r = –0,96) указывает на доминирование аэробного дыхания, тогда как положительная корреляция между CO₂ и H₂ (r = 0,53) отражает вовлечение восстановительных и ферментативных путей в условиях истощения кислорода. Изменения уровней CO₂ и O₂ отражают активность митохондриального дыхания и цикла Кребса — основного пути окисления органических соединений для получения энергии в митохондриях [32]. Солевой стресс нарушает эти процессы, вызывая осмотические и ионные повреждения, ингибируя митохондриальное дыхание и снижая активность ключевых ферментов и связанной с ними дыхательной электронтранспортной цепи, которая регенерирует NAD⁺ для непрерывной работы цикла [35]. Это представляет собой адаптивный ответ, включающий переход в «энергосберегающий режим». Обработка ЭМП в этом контексте проявила двойную роль — стимулирующую (эустресс) в оптимальных условиях и защитную (компенсация дистресса) в условиях солевого стресса.
Особый интерес представляет динамика H₂ как индикатора окислительно-восстановительного дисбаланса и адаптивных реакций у прорастающих семян гороха, где высокая эмиссия H₂ напрямую не связана с циклом Кребса и указывает на активацию гидрогеназных путей, характерных для анаэробиоза. Было показано, что состояние гипоксии во время набухания семян гороха не является редким явлением и может быть обнаружено по уровню фосфоресценции порфиринов [42,43] или по динамике эмиссии H₂ [29]. Его возникновение как неспецифическая реакция на стрессовые факторы в низких дозах наблюдалось во время погружения в воду [29], естественного и ускоренного старения [42], а также после γ-облучения, импульсной световой обработки, облучения гелий-неоновым лазером и воздействия нетермального ЭМП GSM-диапазона (905 МГц) [44] во время прорастания семян в стандартных условиях (без дефицита O₂ в среде). По мнению авторов, гипоксия под семенной оболочкой набухающего семени возникает в результате дисбаланса между активным поглощением O₂ зародышем и его медленной диффузией через семенную оболочку. Также обсуждалась важная роль факторов, влияющих на состояние аквапориновых каналов в плазматических мембранах, отвечающих за скорость поглощения воды и метаболические процессы, а также вовлечение неферментативного гидролиза углеводов и амино-карбонильных реакций в возникновение гипоксии у семян гороха разного качества [43,44].
С другой стороны, известно, что в условиях анаэробиоза растения могут продуцировать водород посредством активации [FeFe]-гидрогеназ в анаэробных условиях, когда дыхательная цепь перегружается и в клетках накапливается NADH. Фермент гидрогеназа окисляет NADH, восстанавливая протоны (H⁺) до молекулярного водорода (H₂). Это представляет собой «аварийный» механизм регенерации NAD⁺ для поддержания необходимого уровня гликолиза [45]; в этом контексте обработка ЭМП может снижать потребность в активации альтернативных путей регенерации NAD⁺, включающих эмиссию H₂ в стрессовых условиях, тем самым поддерживая митохондриальное дыхание [46]. В нашем эксперименте высокая эмиссия H₂ в варианте «Почва + Семена», особенно после обработки ЭМП, может также указывать на то, что в результате активации дыхания и последующего падения концентрации кислорода в закрытой системе в набухающих семенах гороха в гипоксических условиях активируется альтернативный путь регенерации NAD⁺. В условиях солевого стресса из-за повреждения мембран и ингибирования дыхания эмиссия H₂ уменьшилась на порядок. Однако значимые трехсторонние взаимодействия, наблюдаемые для H₂, позволяют предположить, что обработка ЭМП способна смягчать солевой стресс, снижая скорость эмиссии H₂ наполовину, что, скорее всего, указывает на сохранение функциональности митохондрий в набухающих семенах гороха и поддержание оптимального баланса между скоростью дыхания и снабжением кислородом в стрессовых условиях.
Таким образом, анализ данных выявил четкие закономерности, указывающие на глубокое влияние предпосевной биоактивации семян низкочастотным неионизирующим ЭМП, генерируемым устройством «ТОР», на их физиолого-биохимическое состояние, особенно в условиях солевого стресса. Ключевое наблюдение заключается в том, что обработка стабилизирует метаболизм, переводя его в энергосберегающий режим, что наиболее четко проявляется в динамике диоксида углерода (CO₂), кислорода (O₂) и водорода (H₂). То есть обработка семян гороха ЭМП сразу после посева в почву может представлять собой многообещающий нехимический метод повышения толерантности растений к солевому стрессу. В этом контексте ЭМП следует рассматривать как модификатор метаболического ответа, а не как универсальный стимулятор, а эмиссию H₂ следует считать наиболее чувствительным индикатором окислительно-восстановительного дисбаланса, индуцированного гипоксией и модулируемого ЭМП.
Двухфазная динамика NH₃ отражает нелинейное и многостадийное биогеохимическое превращение азота в почве, где баланс между генерацией, потреблением и потерями аммиака определяется состоянием биоты и условиями окружающей среды. Примечательно, что все эти процессы менее чувствительны к электромагнитному воздействию, чем дыхательный и ферментативный метаболизм прорастающих семян. Увеличение концентрации NH₃ на первом этапе (дни 1–2), по-видимому, связано с начальной активацией почвенных микроорганизмов и деградацией белковых структур семян во время прорастания, что приводит к высвобождению аминокислот и последующей аммонификации при протеолизе запасных белков. Этот процесс был наиболее выражен в варианте «Почва + NaCl + Семена», вероятно, как реакция на осмотический шок. Снижение, наблюдаемое к 3-му дню, может быть связано с временным подавлением микробной активности из-за накопления метаболических побочных продуктов, потреблением аммиака самими прорастающими семенами в качестве источника азота для синтеза белка de novo и адсорбцией NH₃ на поверхности почвенных частиц. Второй пик на 4-й день, скорее всего, связан с реактивацией аммонифицирующих бактерий после преодоления временного стресса и разрушением остаточных органических соединений в развивающихся проростках, особенно временных азоттранспортных соединений (например, аспарагина). В этом случае присутствие NaCl дополнительно усиливает этот процесс, проявляясь в более выраженных пиках NH₃.
Согласно литературным данным о влиянии ЭМП на метаболизм растений, следующие биофизические и биохимические механизмы, вероятно, лежат в основе наблюдаемых ранних эффектов ЭМП-прайминга семян гороха:
- Воздействие на мембраны: Низкоинтенсивное ЭМП может модулировать проницаемость клеточных и митохондриальных мембран, тем самым изменяя внутриклеточный ионный баланс и влияя на метаболические процессы, особенно в условиях солевого стресса. Это может служить сигналом для активации адаптивных каскадов [7,47].
- Модуляция окислительно-восстановительного статуса: ЭМП может влиять на работу электронтранспортных цепей в митохондриях семян, изменяя эффективность окислительного фосфорилирования и смещая баланс между аэробными и анаболическими процессами в сторону повышения энергоэффективности дыхательного метаболизма [12–14]. Это снижает генерацию активных форм кислорода (АФК) в стрессовых условиях за счет ингибирования гидрогеназ и перераспределения потока электронов при анаэробном метаболизме [48], что согласуется с наблюдаемым снижением «индуцированного стрессом» дыхания (CO₂) и эмиссии H₂.
- Индукция защитных систем: Косвенно, через окислительно-восстановительную сигнализацию, ЭМП может усиливать экспрессию генов и активность ферментов антиоксидантной защиты и осморегуляции, тем самым повышая общую толерантность проростков [49].
- Косвенные эффекты через почвенный микробиом: Об этом свидетельствуют изменения уровней O₂ и CO₂ в системах без семян, которые впоследствии могут влиять на метаболизм корней [50].
Засоление почвы оказывает выраженное негативное влияние на рост и продуктивность сельскохозяйственных культур, особенно в полузасушливых и засушливых регионах мира, которые составляют примерно одну треть поверхности суши. По данным ФАО (Продовольственная и сельскохозяйственная организация), более 800 миллионов гектаров пахотных земель в мире подвержены различным формам засоления, что представляет собой одну из самых серьезных угроз для глобальной продовольственной безопасности [30,51–54]. Обработку ЭМП можно рассматривать как инструмент физического прайминга семян, способный усиливать дыхание и регенеративные процессы в прорастающих семенах, в том числе в стрессовых соленых условиях, что важно для развития агроинженерных технологий, направленных на управление ростом сельскохозяйственных культур и их стрессоустойчивостью. Полученные данные об изменениях скоростей процессов газообмена создают важную основу для целенаправленных биохимических исследований конкретных ферментативных систем, которые могут служить первичными мишенями для электромагнитного воздействия.
5. Выводы
Было установлено, что предпосевная обработка семян гороха ЭМП проявляет адаптогенные свойства в условиях засоления, способствуя стабилизации аэробного дыхания и снижая признаки метаболического стресса; это наиболее четко отразилось в динамике H₂ и CO₂. ЭМП и засоление оказывают значимые модулирующие эффекты, причем их взаимодействие носит антагонистический характер. Стрессовый фактор ингибировал этот процесс, снижая концентрацию CO₂ к концу эксперимента на 20–30% по сравнению с незасоленными вариантами. Обработка ЭМП в оптимальных условиях стимулировала дыхание, увеличивая концентрацию CO₂ на 15%, тогда как при засолении она, напротив, приводила к снижению на 8–10% относительно необработанного контроля.
Выделение молекулярного водорода (H₂) идентифицировано как высокочувствительный биохимический маркер для оценки эффективности биофизических обработок. В отсутствие солевого стресса обработка ЭМП увеличила выделение H₂ в три раза и сместила его пик на один день раньше, что может указывать на ускоренную активацию защитных систем семян при развивающейся гипоксии. Засоление снизило уровни концентрации H₂ на порядок, в то время как обработка ЭМП стабилизировала скорость его выделения, уменьшив ее почти вдвое. Таким образом, ЭМП следует рассматривать как модификатор метаболического ответа семян на условия набухания, а не только как стимулятор прорастания.
Вклад авторов: Концептуализация, И.М.К.; методология, программное обеспечение, А.С.Ш.; валидация, Е.Г.К. и В.Г.З.; формальный анализ, П.Н.Б.; ресурсы, курирование данных, В.Г.Г.; подготовка первоначального проекта текста, Е.А.Г.; написание текста (рецензирование и редактирование), И.М.К.; руководство, С.Ю.К.; написание текста (рецензирование и редактирование), администрирование проекта, И.Ф.Т.; руководство, методология, привлечение финансирования, Е.В.Б. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование: Это исследование было проведено с использованием средств АО «Концерн «Гранит».
Заявление Этического комитета: Не применимо.
Заявление об информированном согласии: Не применимо.
Заявление о доступности данных: Оригинальные данные, представленные в этом исследовании, включены в статью. Дополнительные запросы можно направлять автору для переписки.
Конфликт интересов: Авторы Игорь Ф. Турканов, Елена В. Бондарчук, Валерий Г. Грязнов и Екатерина А. Галкина были трудоустроены в компании АО «Концерн «Гранит». Остальные авторы заявляют, что исследование было проведено при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Сокращения
В этой рукописи используются следующие сокращения:
| Сокращение | Расшифровка |
|---|---|
| ЭМП | Электромагнитные поля |
| NADH | Никотинамидадениндинуклеотид (восстановленная форма) |
| NAD⁺ | Никотинамидадениндинуклеотид (окисленная форма) |
| RQ | Дыхательный коэффициент |
| ROS | Активные формы кислорода (АФК) |
Литература
- Corbineau, F.; Taskiran-Özbingöl, N.; El-Maarouf-Bouteau, H. Улучшение качества семян с помощью прайминга: концепция и биологические основы. Seeds [Семена] 2023, *2*, 101–115. [CrossRef]
- De Souza, A.; García, D.; Sueiro, L.; Gilart, F. Предпосевная магнитная обработка семян томатов увеличивает рост и урожайность растений. Bioelectromagnetics [Биоэлектромагнетизм] 2006, *27*, 247–257. [CrossRef]
- Vian, A.; Davies, E.; Gendraud, M.; Bonnet, P. Реакции растений на высокочастотные электромагнитные поля. BioMed Res. Int. [Биомедицинские исследования, международное издание] 2016, *2016*, 1830262. [CrossRef]
- Galland, P.; Pazur, A. Магниторецепция у растений. J. Plant Res. [Журнал исследований растений] 2005, *118*, 371–389. [CrossRef] [PubMed]
- Maffei, M.E. Влияние магнитного поля на рост, развитие и эволюцию растений. Front. Plant Sci. [Границы науки о растениях] 2014, *5*, 445. [CrossRef]
- Abdulraheem, M.I.; Moshood, A.Y.; Gourkhede, P.H.; Li, L.; Zhang, Y.; Abdi, G.; Raghavan, V.; Hu, J. Новые тенденции в применении импульсных магнитных полей для модуляции роста и развития растений: механизмы, динамика и потенциальное влияние. Acta Physiol. Plant. [Физиология растений] 2025, *47*, 94. [CrossRef]
- Belyavskaya, N.A. Биологические эффекты слабого магнитного поля на растения. Adv. Space Res. [Достижения в космических исследованиях] 2004, *34*, 1566–1574. [CrossRef]
- Turkanov, I.F.; Bondarchuk, E.V.; Gryaznov, V.G.; Galkina, E.A.; Guzenko, A.Y.; Zainullin, V.G.; Kozar, E.G.; Kaigorodova, I.M. Эффекты дистанционной обработки семян ячменя слабыми нетермальными импульсными электромагнитными полями на развитие растений и урожайность. Seeds [Семена] 2025, *4*, 35. [CrossRef]
- Qin, Y.; Ma, R.; Wang, Z.; Liu, Z.; Chen, L.; Han, Q.; Liu, Z.; Liu, G. Влияние солевого стресса на прорастание семян и рост зародышей Oxytropis coerulea. Legume Res. Int. J. [Зернобобовые исследования, международный журнал] 2025, *48*, 38–46. [CrossRef]
- Ntshalintshali, S.; Sibiya, S.G.P.; Mabuza, N.; Nkomo, M.; Buthelezi, L.G. Прайминг семян зернобобовых культур: механизмы, применение и будущие направления для улучшения прорастания, роста и устойчивости к абиотическим стрессам в устойчивом сельском хозяйстве: обзор. J. Agron. Crop Sci. [Журнал агрономии и науки о растениях] 2025, *211*, e70115. [CrossRef]
- Shabala, S.; Bose, J.; Hedrich, R. Солевые пузыри: имеют ли они значение? Trends Plant Sci. [Тенденции в науке о растениях] 2014, *19*, 687–691. [CrossRef]
- Cakmak, T.; Dumlupinar, R.; Erdal, S. Ускорение прорастания и раннего роста проростков пшеницы и фасоли, выращенных в различных магнитных и осмотических условиях. Bioelectromagnetics [Биоэлектромагнетизм] 2010, *31*, 120–129. [CrossRef]
- Vashisth, A.; Nagarajan, S. Влияние статического магнитного поля на прорастание и характеристики раннего роста семян подсолнечника (Helianthus annuus). J. Plant Physiol. [Журнал физиологии растений] 2010, *167*, 149–156. [CrossRef]
- Van Aken, O. Митохондриальные окислительно-восстановительные системы как центральные узлы в метаболизме и сигнализации растений. Plant Physiol. [Физиология растений] 2021, *186*, 36–52. [CrossRef]
- Wang, L.; Zuo, Q.; Zheng, J.; You, J.; Yang, G.; Leng, S. Солевой стресс снижает урожайность семян и замедляет процесс роста рапса (Brassica napus L.) за счет изменения азотного и углеродного статуса. Sci. Rep. [Научные доклады] 2022, *12*, 17884. [CrossRef] [PubMed]
- Waheed, A.; Zhuo, L.; Wang, M.; Hailiang, X.; Tong, Z.; Wang, C.; Aili, A. Интегративные механизмы солеустойчивости растений: биологические пути, фитогормональная регуляция и технологические инновации. Plant Stress [Стресс растений] 2024, *14*, 100652. [CrossRef]
- Ben Youssef, R.; Jelali, N.; Acosta Motos, J.R.; Abdelly, C.; Albacete, A. Прайминг семян салициловой кислотой: ключевой рубеж в обеспечении толерантности к солевому стрессу при прорастании семян ячменя и росте проростков. Agronomy [Агрономия] 2025, *15*, 154. [CrossRef]
- Shrivastava, P.; Kumar, R. Засоление почв: серьезная экологическая проблема и бактерии, стимулирующие рост растений, как один из инструментов для ее смягчения. Saudi J. Biol. Sci. [Саудовский журнал биологических наук] 2015, *22*, 123–131. [CrossRef] [PubMed]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.-K.; Shabala, S. Механизмы ответа растений и адаптации к засолению почв. Innovation [Инновации] 2020, *1*, 100017. [CrossRef]
- Sharafi, S. Повышение устойчивости проростков люцерны улитковой к стрессу засоления-засухи с использованием магнитного поля и ультразвуковых волн. Greenh. Plant Prod. J. [Журнал тепличного растениеводства] 2024, *1*, 27–37. [CrossRef]
- Evrendilek, G.A.; Yalçın, B. Повышение качества и устойчивости семян томатов с помощью импульсных электрических полей: улучшение прорастания, стрессоустойчивости и микробной дезинфекции. Appl. Sci. [Прикладные науки] 2025, *15*, 6447. [CrossRef]
- Sharafi, S. Улучшенный рост проростков однолетнего эспарцета в условиях соле-засушливого стресса с помощью обработки ультразвуком и магнитным полем. Appl. Water Sci. [Прикладные водные науки] 2025, *15*, 59. [CrossRef]
- Cesniene, I.; Cesna, V.; Miskelyte, D.; Novickij, V.; Mildaziene, V.; Sirgedaite-Seziene, V. Обработка семян холодной плазмой и электромагнитным полем: изменения антиоксидантной способности проростков у разных полусибсовых семейств Picea abies (L.) H. Karst. Plants [Растения] 2024, *13*, 2021. [CrossRef] [PubMed]
- Bhavani, L.L.; Dayal, A. Влияние магнитной и электрической обработки на рост, урожайность и параметры проростков нута (Cicer arietinum L.). Environ. Conserv. J. [Журнал охраны окружающей среды] 2024, *25*, 726–733. [CrossRef]
- Mandal, A.; Bala, N. Влияние электротерапии и солевого прайминга на улучшение всхожести и энергии прорастания семян пшеницы. J. Soils Crops [Журнал почв и сельскохозяйственных культур] 2023, *32*, 317–323. [CrossRef]
- Aleman, E.I.; Diaz, R.S.; Dubois, A.F.; Boix, Y.F.; Aguilera, J.G.; Zuffo, A.M.; Steiner, F. Влияние неоднородных электромагнитных полей частотой 60 Гц на прорастание семян томатов (сорт L-05), фотосинтез и рост проростков в условиях солевого стресса. Bioelectromagnetics [Биоэлектромагнетизм] 2025, *46*, e22537. [CrossRef]
- Mumtaz, S.; Javed, R.; Rana, J.N.; Iqbal, M.; Choi, E.H. Прайминг семян импульсным мощным СВЧ-излучением модулирует прорастание, рост, окислительно-восстановительный гомеостаз и гормональные сдвиги у ячменя для улучшения роста проростков: раскрытие молекулярной динамики. Free. Radic. Biol. Med. [Свободные радикалы в биологии и медицине] 2024, *222*, 371–385. [CrossRef]
- Kataria, S.; Jain, M.; Tripathi, D.K.; Singh, V.P. Участие продукции оксида азота, зависимой от нитратредуктазы, в индуцированной магнитным праймингом солеустойчивости сои. Physiol. Plant. [Физиология растений] 2020, *168*, 422–436. [CrossRef] [PubMed]
- Khashirova, S.Y.; Shabaev, A.S.; Bondarchuk, E.V.; Turkanov, I.F.; Gryaznov, V.G.; Galkina, E.A.; Kaigorodova, I.M.; Zainullin, V.G. Газохроматографический анализ сельскохозяйственных почв и семян, обработанных слабыми неионизирующими нетермальными электромагнитными полями (ЭМП). Известия Коми научного центра УрО РАН. Серия «Сельскохозяйственные науки» 2025, *1*, 120–126. (На русском) [CrossRef]
- Sarraf, M.; Kataria, S.; Taimourya, H.; Santos, L.O.; Menegatti, R.D.; Jain, M.; Ihtisham, M.; Liu, S. Применение магнитного поля (МП) у растений: обзор. Plants [Растения] 2020, *9*, 1139. [CrossRef]
- Tomaz, A.; Palma, P.; Alvarenga, P.; Gonçalves, M.C. Глава 10 — Риск засоления почв в условиях изменения климата и его влияние на здоровье почвы и урожайность сельскохозяйственных культур. В книге Climate Change and Soil Interactions [Изменение климата и взаимодействие с почвой]; Elsevier: Амстердам, Нидерланды, 2026; стр. 235–272. [CrossRef]
- Tcherkez, G.; Ghashghaie, J. Plant Respiration: Metabolic Fluxes and Carbon Balance [Дыхание растений: метаболические потоки и углеродный баланс]; Springer International Publishing: Берлин/Гейдельберг, Германия, 2017; Том 334.
- Russell, G.; Zulfiqar, F.; Hancock, J.T. Гидрогеназы и роль молекулярного водорода у растений. Plants [Растения] 2020, *9*, 1136. [CrossRef] [PubMed]
- Zhao, G.; Cheng, P.; Zhang, T.; Abdalmegeed, D.; Xu, S.; Shen, W. Вода, обогащенная водородом, приготовленная с использованием аммиак-борана, может повышать устойчивость проростков рапса (Brassica napus L.) к засолению, засухе или кадмию. Ecotoxicol. Environ. Saf. [Экотоксикология и экологическая безопасность] 2021, *224*, 112640. [CrossRef] [PubMed]
- Dhal, S.; Pal, H. Регуляция гликолиза и цикла Кребса во время биотических и абиотических стрессов. В книге Photosynthesis and Respiratory Cycles During Environmental Stress Response in Plants [Фотосинтез и дыхательные циклы при реакции растений на экологический стресс]; Apple Academic Press: Палм-Бэй, Флорида, США, 2022; стр. 263–308.
- Jin, Q.; Zhu, K.; Cui, W.; Xie, Y.; Han, B.; Shen, W. Газообразный водород действует как новая биоактивная молекула, повышая толерантность растений к окислительному стрессу, индуцированному паракватом, через модуляцию сигнальной системы гемоксигеназы-1. Plant Cell Environ. [Окружающая среда растительной клетки] 2013, *36*, 956–969. [CrossRef]
- Xu, S.; Zhu, S.; Jiang, Y.; Wang, N.; Wang, R.; Shen, W.; Yang, J. Вода, обогащенная водородом, снижает солевой стресс у риса во время прорастания семян. Plant Soil [Почва растений] 2013, *370*, 47–57. [CrossRef]
- Fu, X.; Ma, L.; Gui, R.; Ashraf, U.; Li, Y.; Yang, X.; Zhang, J.; Imran, M.; Tang, X.; Tiana, H.; и др. Дифференциальная реакция ароматных сортов риса на засоление и воду, обогащенную водородом, в связи с ростом и антиоксидантными защитными механизмами. Int. J. Phytoremediation [Международный журнал фиторемедиации] 2021, *23*, 1203–1211. [CrossRef]
- Tarchokov, H.S.; Chochaev, M.M.; Mataeva, O.K.; Shogenov, A.K.; Kushkhabiev, A.Z. Влияние способов посева сельскохозяйственных культур на интенсивность эрозионных процессов и урожайность на склоновых землях Кабардино-Балкарской Республики. Известия Кабардино-Балкарского научного центра РАН 2020, *5*, 5–19. (На русском) [CrossRef]
- Самохвалов, С.Г.; Прижукова, В.Г.; Молканова, Л.И. Методические указания по определению щелочногидролизуемого азота в почве по методу Корнфилда; Издательство ЦИНАО: Москва, Россия, 1985; 8 с. (На русском)
- Duncan, D.B. Множественный диапазонный и множественный F-критерии. Biometrics [Биометрия] 1955, *11*, 1–42. [CrossRef]
- Veselova, T.V.; Veselovskii, V.A.; Usmanov, P.D.; Usmanova, O.V.; Kozar, V.I. Гипоксия и повреждения при набухании стареющих семян. Russ. J. Plant Physiol. [Российский журнал физиологии растений] 2003, *50*, 835–842. [CrossRef]
- Veselova, T.V.; Veselovsky, V.A.; Turovetsky, V.B.; Galchuk, S.V.; Vanyushin, B.F.; Aleksandrushkina, N.I.; Rubin, A.B. Постгипоксический окислительный стресс после появления корешка как возможная причина появления аномальных проростков гороха. Seed Sci. Technol. [Наука и технология семян] 2004, *32*, 283–296. [CrossRef]
- Veselova, T.V.; Veselovsky, V.A.; Deev, L.I.; Baizhumanov, A.A. Нетепловой эффект электромагнитного излучения GSM-диапазона на качество семян гороха. Радиационная биология. Радиоэкология 2012, *52*, 394.
- Maric, A.K.; Radan, A.; Milcic, N.; Sudar, M.; Findrik Blazevic, Z. Гидрогеназы — типы, источники, свойства и потенциал их применения. Kem. Ind. Cas. Kem. Kem. Inz. Hrvat. [Химическая промышленность] 2025, *74*, 33–42. [CrossRef]
- Chow, S.C.; Zhang, Y.; Ng, R.W.; Hui, S.Y.R.; Solovyov, I.A.; Lui, W.Y. Внешнее радиочастотное ЭМП изменяет количество клеток и баланс АФК, возможно, посредством регуляции метаболизма NADPH и апоптоза. Front. Public Health [Границы общественного здравоохранения] 2024, *12*, 1425023. [CrossRef]
- Pietruszewski, S.; Martínez, E. Магнитное поле как метод улучшения качества посевного материала: обзор. Int. Agrophys. [Международная агрофизика] 2015, *29*, 377–389. [CrossRef]
- Stiebritz, M.T.; Reiher, M. Гидрогеназы и кислород. Chem. Sci. [Химические науки] 2012, *3*, 1739–1751. [CrossRef]
- Chaudhary, D.; Aggarwal, H.; Kaur, B.; Agarwal, H.; Das, A.; Kumar, A.; Mishra, V.; Kumar Singh, P.; Joshi, N.C. Прайминг — состояние готовности к экологическим стрессам у растений. Plant Gene [Ген растений] 2025, *43*, 100513. [CrossRef]
- Suvendran, S.; Johnson, D.; Acevedo, M.; Smithers, B.; Xu, P. Электромагнитная обработка воды и внесение почвенного компоста для снижения последствий засоления почв. Water [Вода] 2024, *16*, 1577. [CrossRef]
- Golla, B. Система сельскохозяйственного производства в засушливых и полузасушливых регионах. Int. J. Agric. Sci. Food Technol. [Международный журнал сельскохозяйственных наук и пищевых технологий] 2021, *7*, 234–244. [CrossRef]
- Aksoy, S.; Yildirim, A.; Gorji, T.; Hamzehpour, N.; Tanik, A.; Sertel, E. Оценка эффективности алгоритмов машинного обучения для картографирования засоления почв на платформе Google Earth Engine с использованием данных Sentinel-2A и Landsat-8 OLI. Adv. Space Res. [Достижения в космических исследованиях] 2022, *69*, 1072–1086. [CrossRef]
- Saxena, N.P.; Saxena, M.C.; Johansen, C.; Virmani, S.M.; Harris, H. Будущие приоритеты исследований нута в регионе Западной Азии и Северной Африки (WANA) и в полузасушливых тропиках (SAT). В сборнике Adaptation of Chickpea in the West Asia and North Africa Region [Адаптация нута в регионе Западной Азии и Северной Африки]; ICARDA: Алеппо, Сирия, 1996; стр. 257–262.
- Продовольственная и сельскохозяйственная организация ООН (ФАО). Доступно онлайн: https://www.fao.org (дата обращения: 20 ноября 2025 г.).
Отказ от ответственности издателя/примечание для издателя: Утверждения, мнения и данные, содержащиеся во всех публикациях, принадлежат исключительно отдельному(ым) автору(ам) и лиц(у), внесшему(им) вклад, и не относятся к MDPI и/или редактору(ам). MDPI и/или редактор(ы) снимают с себя ответственность за любой вред, причиненный людям или имуществу, возникший в результате использования любых идей, методов, инструкций или продуктов, упомянутых в содержани